
Abstract
Introduction
The term proprioception refers to the perception of the locomotor system’s condition, based on the signals originating in the locomotor system itself (Sherrington, 1909). This information is essential for motor control: deafferented patients are not only unable to control their gait and movements in the absence of visual information, but they also show remarkable deficits even when visual information is available (Gallagher, 2006; Sainburg et al., 1993). Proprioceptive input underlies various types of sensations, for example, it is the basis of the sense of joint position, movement, and force (Proske & Gandevia, 2012). In this study, we focused on joint position sense.
People with intact proprioception show considerable individual differences with respect to proprioceptive acuity and the ability to perceive proprioceptive stimuli (Han et al., 2016). These skills are associated with physical accomplishment: athletes with higher accuracy tend to achieve higher level of performance (Han et al., 2015) and get less likely injured (Cameron et al., 2003; Payne et al., 1997). To assess proprioceptive acuity, one of the most widely used tests is Joint Position Reproduction (JPR) test (Han et al., 2016). In the ipsilateral version of the JRP test, one limb of the participant is moved from starting position to a target position then it is moved back to the starting position. From here, participants have to reproduce the target position with the same limb as accurately as possible. The procedure requires the storage of the target position in short-term memory (Goble, 2010).
Goble, Mousigian, and Brown (2012) demonstrated that working memory capacity influences performance in JPR test, if participants are asked to perform a simultaneous task (i.e., counting upwards by three from a random number between 1 and 100). In their experiment, elderly participants with low working memory capacity showed decreased proprioceptive acuity while conducting a competing task. Those with high capacity showed no decline in performance though. In line with these findings, Boisgontier and colleagues (2012) demonstrated that proprioceptive processing becomes cognitively more demanding in older age. They also compared the performance of elderly and young individuals on JPR test, while doing a secondary task simultaneously. The performance of elderly people decreased to a greater extent when they had to do a secondary, cognitively demanding task (i.e., Stroop Task). Also, Alloway and Alloway (2015) found that a proprioceptively demanding training can increase working memory capacity, which supports the idea that proprioceptive processing requires working memory resources. Finally, Yasuda and colleagues (2014) reported that attentional load has an impact on proprioceptive accuracy. In their experiment, the performance of healthy young participants was influenced negatively only by a high cognitive demand exercise (i.e., subtracting 7 from random numbers), but not by a low cognitive demand exercise (i.e., subtracting 3 from random numbers).
As working memory is a multi-component system, traditionally encompassing two storage subsystems, that is, the visuospatial sketchpad and the phonological loop (Baddeley & Hitch, 1974), a fundamental question is in which subsystem proprioceptive information is stored. To our knowledge, only one study has investigated the modality-specific storage of proprioceptive information to date. Goble, Aaron, and colleagues (2012) found that patients with cerebral palsy, as well as healthy adults (Goble et al., 2010), were more accurate when they had more time (15 s) to encode joint positions than when processing time was limited (2 s). However, the patients’ improvement in proprioceptive accuracy in the longer presentation time condition was positively associated with spatial short-term memory capacity at a moderate to strong level (
To test the role of working memory subsystems in storing proprioceptive information, we developed and used a modified version of the JPR test. In this test, as in other working memory measurements, the ability to retain more than one element (joint position) at one time was assessed and a capacity score for every participant was calculated. To test the paradigm, we recruited a sample of healthy individuals without any neurological or psychiatric disorders. Based on Goble, Aaron, and colleagues’ (2012) work, it was hypothesised that joint positions are stored in the visuospatial sketchpad of working memory, thus the capacity to retain multiple proprioceptively determined joint positions will show a positive association with spatial capacity (Hypothesis 1), and no association with verbal capacity (Hypothesis 2).
Methods
Participants
Sample size calculation for a medium level correlation (
Measurements
Single-position error
We used the ipsilateral version of the JPR test and measured single-position error in the elbow joint of the non-dominant hand. We chose to measure non-dominant hand, because it was found to be more accurate than the dominant hand in proprioceptive processing (Goble & Brown, 2008). Measurements were made using a device similar to that described by Goble (2010). The device allowed the rotation of the elbow joint by moving the lower arm while the upper arm and the shoulder remained in a stable position. The 180 degree indicated fully extended elbow, and approximately 10–15 degree meant fully flexed elbow. Starting position was always 180 degree. The arm of the participant was moved from the starting position to one of the target positions, and kept there for 4 s. After that, the arm was moved back to the starting position, and the participants were asked to reproduce the target position by actively moving her or his arm. We used nine target positions (30, 45, 60, 75, 90, 105, 120, 135, and 150 degree), and presented them in random order. To determine the persons’ single-position error score, we took the absolute value of the nine error scores (i.e., the difference between the presented and reproduced angle in degrees) and averaged them.
Proprioceptive short-term memory task
We applied a modified version of the above described ipsilateral JPR test to assess proprioceptive short-term memory. We used the same nine target positions as in the single-position task, from 30 to 150 degree. We composed quasi-random sequences of positions, that is, one position was not included multiple times in one sequence. The test started with the presentation and reproduction of a sequence of two joint positions. In every trial, we increased the number of the presented positions by one. Independently from their performance, each participant executed the complete test, ending with the reproduction of an 8-position sequence. We repeated this procedure (presenting 2–8 joint positions) three times overall, so all of the participants were presented with 3 sequences in each length, 21 sequences in total. Participants always had to reproduce the positions in the same order as they were presented. The absolute value of the difference between the presented and the reproduced position was considered the error score for the given position.
In the first step, similar to other working memory measurements, we intended to determine a proprioceptive memory capacity score for every person. In other words, we attempted to find the maximal number of positions which a person can retrieve. In classical short-term memory measurements, such as the Digit span task or the Corsi block task, one will get the capacity score as the number of the elements in the longest sequence which the participants can retrieve two times correctly out of three attempts. However, in the case of proprioception, determining the correctness of a sequence is difficult as the error score is a continuous value. To solve this problem, in the first step, we calculated the mean of the absolute errors with respect to the 21 sequences. Figure 1 shows an example of the scoring of one sequence containing 3 elements.

Example of the establishment of the error score in the case of a sequence, containing 3 positions.
In the next step of the analysis, error scores for each sequence length (2–8) in each trial (1-3) were calculated. We hypothesised that a significant drop in performance occurs after reaching the proprioceptive capacity. Thus, we wanted to identify the sequence after which the magnitude of the errors starts to disproportionately increase. To this end, we used a modified step function of the “lsqcurvefit” function of Matlab (The MathWorks Inc., Natick, MA, USA). To be able to handle non-continuous data, the step function was modified by drawing a 1-unit long transition line around the breakpoint. The curve was fitted to achieve the least sum of the squares for errors (SSE). Proprioceptive capacity was determined as the middle point of the transition line (showing the “breakpoint”) (see Figure 2 for an example).

Example of the determination of proprioceptive memory capacity. In this case, proprioceptive memory capacity was determined as 4.40. . Circles represent the individual error score of a given sequence, and squares represent the average error score for the respective sequence length. The solid line represents the step function, and the vertical dashed line shows the middle point of the transition line (proprioceptive capacity). The dotted line represents the linear curve, which was fitted to compare goodness of fit values.
By calculating the mean of the three error scores belonging to the same length sequence, we also calculated one error score with respect to every sequence length (2–8 position error) for a further analysis (see Table 1).
Descriptive statistics of the measured variables.

Verbal short-term memory capacity
We used the Digit span task of PEBL software (version 0.14) (Mueller, 2014) to assess verbal capacity. In this task, participants have to memorise sequences of randomly presented digits, and reproduce them in the same order as presented, by typing them with the keyboard of the computer. The first sequence contains three numbers. If one reproduces the sequence correctly, the length of the next sequence increases by one. The test ends, when participant gives two times incorrect answer to a particular sequence length. Verbal capacity is determined by the number of elements in the longest correctly reproduced sequence.
Spatial short-term memory capacity
We used the Corsi blocks task (Corsi, 1972) of PEBL software (version 0.14) (Mueller, 2014) to assess spatial capacity. In this task, participants have to memorise spatial locations. There are 9 possible locations, appearing as blue squares on the computer screen. The squares flash up in a random order following each other. Participants have to memorise and reproduce the sequence of the squares in the same order as presented by clicking on the squares with the mouse button. The test starts with two locations to memorise, and the number of the presented locations increases by one if the participant can correctly reproduce the sequence. The test ends, when participant reproduces incorrectly two times a particular sequence length. Spatial capacity is determined by the number of the elements in the longest correctly reproduced sequence.
Protocol
The order of the tasks was the following for all participants: Single-position error, Proprioceptive Short-term memory task, Digit span task, Corsi block task. During the proprioceptive measurements, participants’ eyes were covered to prevent the use of visual information.
Results
Table 1 presents the descriptive statistics of the variables. According to Shapiro–Wilk test, the majority of the assessed variables showed a non-normal distribution thus Kendall’s correlation was used to estimate the association between them.
We used 0.9.0.1 version of JASP (JASP Team, 2018) and R software for the statistical analysis.
In the first step of the analysis, we evaluated the goodness of fit of the modified step function, which was used to determine proprioceptive capacity. A linear curve, which minimised the SSE (linear regression), was also fitted on the error scores for each individual. In this way, we were able to evaluate the goodness of fit of the step function against the linear function by comparing the SSE values. SSE was smaller for the step function than for the linear function for 29 participants (74%). When statistically comparing the values with Wilcoxon signed-rank test, a significant difference strongly favouring the unit step function was found (
In the next step, we statistically compared the error scores of the different sequence length with Friedman test, and found a significant difference, χ2(6) = 156.726,
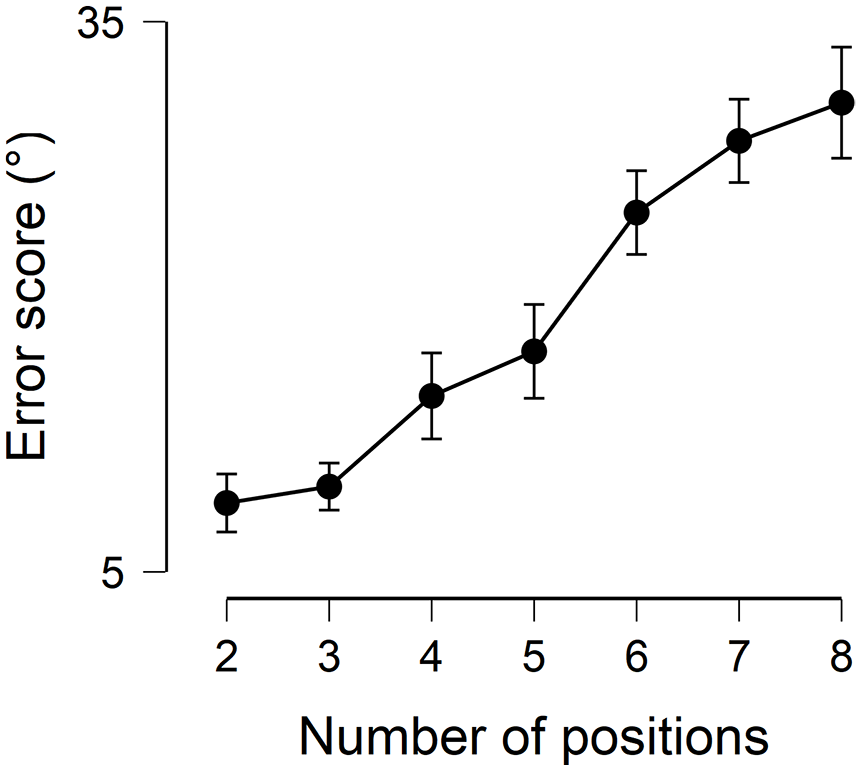
Horizontal axis: number of retained positions, vertical axis: mean error score across participants, error bar: 95 confidence interval.
To test our hypotheses, we conducted correlation analysis (Kendall’s tau) using both the Bayesian and frequentist approaches. For the Bayesian analysis, we used default priors in JASP software. We used two-tailed significance levels, and to reveal the association between Spatial capacity and proprioceptive capacity one-tailed significance test for positive correlation was also used in accordance with our main hypothesis. With two-tailed significance level, frequentist approach did not support the rejection of the null hypothesis in any of the cases (
Associations (Kendall’s tau correlation coefficients, their significance values, and Bayes factors) between the assessed variables.

One-tailed

Scatter plots of the investigated relationships.
Overall, we did not find evidence that would support the assumed association between Proprioceptive short-term memory and Spatial (Hypothesis 1) or Verbal (Hypothesis 2) memory span. Moreover, we found evidence which supports the independence of them (with two-tailed tests). The one-tailed test did not provide convincing evidence for the associations either.
In a subsequent analysis, we also investigated the associations between performance on sequences of different length (2-8 position error) with respect to verbal, spatial, and proprioceptive capacity (Table 3). Note that proprioceptive capacity was derived from 2 to 8 position error with a curve fitting method, so the correlation might be arbitrary here. However, we included this variable in the analysis too, to explore at which sequence length proprioceptive capacity intervene performance. For the
Associations (Kendall’s tau correlation coefficients) of spatial, verbal, and proprioceptive memory capacities with average errors of different sequence lengths in the proprioceptive task. Bonferroni corrected


Scatter plots of the supported correlations between memory capacities (spatial, verbal and proprioceptive) and average errors of different sequence lengths in the proprioceptive memory task.
Finally, it is possible that the relationship between spatial or verbal span and error shown in sequences of different length depends on the level of Proprioceptive span. So we also conducted a series of moderation analyses. The moderator variable was always the proprioceptive span, whereas the predictor variable was the Verbal or the Spatial span, and the outcome variable was 2, 3, 4, 5, 6, 7, or 8 position error. The moderation term was significant for one case: Proprioceptive span was found to be a moderator (
Discussion
In this study, we have developed and used a new procedure to measure proprioceptive short-term memory. In this procedure, participants had to memorise and reproduce an increasing number of proprioceptively determined joint positions. Based on the drop in performance, we derived a proprioceptive capacity score for every individual. Contrary to our primary hypothesis, proprioceptive short-term memory capacity did not show a significant correlation with spatial short-term memory capacity in young individuals. This finding does not support the previous assumption (Goble, Aaron, et al., 2012), that is, that the visuospatial sketchpad of working memory plays a fundamental role in the storage of proprioceptive information. Proprioceptive capacity was not associated with verbal capacity either, which indicates that proprioceptive information is not stored in the phonological loop. Moreover, the independence of proprioceptive span and single-position proprioceptive error was also revealed, which shows that the reproduction accuracy of a single joint position is not related to the capacity to store multiple proprioceptively determined joint positions in working memory.
In a subsequent analysis, we broke down the performance shown in proprioceptive memory task for different sequence lengths and found that proprioceptive memory capacity correlates positively with the performance only if 5 elements have to be replicated, and verbal capacity has a positive association with the performance if 7 elements have to be replicated. Moreover, based on the Bayesian analysis, we can assume that it is also probable that verbal capacity correlates positively with the proprioceptive performance for 6 positions, and spatial capacity in the case of 5 position sequences. It was also revealed that the association between 5 position error and spatial span was only present if Proprioceptive span of the participant was low.
Overall, these findings indicate the existence of a proprioceptive short-term memory span, which is largely independent of spatial and verbal spans, and generally plays a substantial role if one has to remember 5 proprioceptively determined joint positions on average. Verbal capacity underlies performance if 7 positions have to be memorised, and it is feasible to assume that it also plays a role for 6 positions. This association implies that verbal strategies, such as labelling might help the retention of proprioceptive information, especially if the number of presented positions exceeds proprioceptive capacity. Spatial capacity plays a role if 5 positions have to be retained, but only if Proprioceptive capacity is low. Thus, spatial coding may also be an alternative form for coding proprioceptively determined joint position, presumably if the number of joint position exceeds the Proprioceptive span.
If there is a short-term proprioceptive memory capacity, which is independent of both the visuospatial sketchpad and phonological loop, then the next question concerns its storage. In the study of Smyth and colleagues (1988), the capacity to reproduce sequences of visually observed, previously practised or non-practised, body-related movements was measured with or without a concurrent verbal or spatial task. They also investigated how motor interference influences spatial or verbal short-term memory performance. On the one hand, it was found that the concurrent spatial task did not interfere with the reproduction of movement sequences, while the verbal task interfered with it. However, this effect disappeared when participants could preliminary practice the elements of the movement sequences. On the other hand, a concurrent motor task disrupted the reproduction of movement, but not that of verbal or spatial sequences. Based on the logic of interference studies (Baddeley, 1992), it was concluded that a distinct subsystem exists in working memory that stores (body-related) movements. Since then, further empirical research confirmed this assumption (Moreau, 2013; Smyth & Pendleton, 1990). It is possible that proprioceptively determined joint positions are stored in working memory in the same way as visually observed movements, that is, in a dedicated subsystem which is responsible for holding motor information. This possibility is also consistent with the observation that people have a tendency to store single joint positions in form of motor commands (Fel’dman & Latash, 1982). In this light, it is also probable that people store a sequence of joint positions as a sequence of motor commands. This assumption would be worthy of further investigation.
The independence of proprioceptive and spatial span is in contraction with the assumption of Goble, Aaron, and colleagues (2012), namely that visuospatial sketchpad plays a substantial role in the storage of proprioceptive information. This assumption is based on the findings of an empirical study, with the participation of patients with cerebral palsy. It was found that the reproduction accuracy of a single joint position increased when positions were presented for relatively long time compared with a relatively short presentation time. The level of increment in acuity was associated with spatial short-term memory capacity of the participants (Goble, Aaron, et al., 2012). To give an explanation for this finding, one has to keep in mind that cerebral palsy is characterised by severe motor deficits (Bax et al., 2005); thus, patients with palsy may encode proprioceptively presented joint position in a different way than healthy individuals. An evidence for such, a relationship was provided by the results of the present study (i.e., that spatial capacity was associated with the performance only if Proprioceptive capacity was low). In addition, in the above mentioned study, the retention accuracy of a single joint position was measured. The ability to retain various joint positions at one time may be a more appropriate method to investigate the storage-specific characteristics of working memory. A further difference between storing one or several joint positions is that if one has to reproduce a single joint position, it can be done with little time passing between the presentation and the reproduction phase. However, when more joint positions need to be reproduced, the time between the presentation and reproduction phase increases, which may change the frame of reference people use to store proprioceptive information (Gaunet & Rossetti, 2006).
Limitations and further directions
The relatively small sample and the special population (university students) limit the generalisability of the findings of the present study. For example, young people process proprioceptive information more accurately than elderly individuals (Adamo et al., 2009). Also, most of the participants were engaged in regular physical activity, and motor expertise can modulate both proprioceptive acuity (Adamo et al., 2009; Goble, 2010; Niespodziński et al., 2018; Ribeiro & Oliveira, 2011) and the processing of movements in working memory (Moreau, 2013).
It is also important to note that in our study, proprioceptive accuracy measures (both single and multi-position) required active motion from the participants to reproduce the joint positions. However, in JPR task, the reproduction can also happen by passively moving the participants’ arm, and there is only a weak association between the two versions (active and passive) of the test (Elangovan et al., 2014). It is possible that joint positions are stored in a different way in working memory if they had to be reproduced by active or passive motion, so our results apply only to the latter version.
Regarding the ecological validity of the findings, in our paradigm participants had to focus on movements of one joint (elbow) and in one plain (vertical). In natural conditions, when people have to attend movements (e.g., learning new motor skills), these movements are more complex, that is, multiple joints and muscles are involved (Tripp et al., 2006). It is possible to measure the reproduction accuracy of more complex movements by using inclinometer, which was found to be a reliable method to measure proprioceptive accuracy (Dover & Powers, 2003). From a practical point of view, another notable point is that in our study the task implied conscious perception of joint positions, and many studies demonstrated that action and perception does not necessary depend on each other (Rossetti, 1998).
Regarding the methodology, a simple correlational approach was applied in the present study. A widely used approach to study storage specific characteristics in working memory is the interference paradigm (Baddeley, 1992). The basic idea of this method is that if two tasks share the same mental resources (e.g., modality-specific subsystem in working memory), then the parallel execution of them will decrease the performance shown in one or both (Baddeley, 1992). To further explore the modality-specific mechanisms of proprioceptive stimuli, it would be necessary to investigate how spatial, visual, verbal, and motor interferences influence proprioceptive memory capacity.
Conclusion
In our study, we developed and used a new method to measure proprioceptive memory capacity. Our results do not support the idea that proprioceptively determined joint positions are stored in the visuospatial sketchpad of working memory. The independence of span measures indicates that proprioceptive information is stored in a subsystem independent of the visuospatial sketchpad or phonological loop.