Brucellosis is a worldwide zoonosis caused by Brucella species and represents a serious threat to both human and animal health. Omp25 is an important immunogenic and protective antigen in Brucella species; however, the functional mechanism of Omp25 in macrophages has not yet been elucidated. Here, we constructed a Brucella melitensis omp25 deletion mutant (M5-90-Δomp25) and performed microRNA (miRNA) profiling of infected RAW264.7 cells. Eight differentially expressed miRNAs (mmu-miR-146a-5p, mmu-miR-155-5p, mmu-miR-3473a, mmu-miR-149-3p, mmu-miR-671-5p, mmu-miR-1224-5p, mmu-miR-1895, and mmu-miR-5126) were identified, with quantitative real-time PCR (qRT-PCR) analysis confirming the up-regulation of mmu-miR-146-a-5p and mmu-miR-155-5p and down-regulation of mmu-miR-149-3p and mmu-miR-5126. mRNA profiling of B. melitensis M5-90-Δomp25-infected RAW264.7 cells identified 967 differentially expressed genes (DEGs) (fold change ≥ 2). Among these, we focused on genes that were predicted by TargetScan, miRanda, and PicTar to be the potential targets of the differentially expressed miRNAs. The results suggested that 17 separate genes are potentially targeted by mmu-miR-149-3p, with one of these genes, Tbr1, also targeted by mmu-miR-5126. qRT-PCR analysis confirmed the up-regulation of nine of the predicted target genes. Our findings provide important information about the functional molecules in host cells, including miRNA and their target genes, affected by Omp25 from Brucella. This information is particularly valuable for the prophylaxis and treatment of brucellosis.
Brucellosis is a worldwide zoonosis and serious threat to the health of humans and animals alike. Brucellosis is caused by Brucella species, including B. abortus, B. melitensis, B. suis, B. ovis, B. neotomae, and B. cannis, all of which show differences in pathogenicity and host preference.1 As intracellular bacterial pathogens, Brucella species invade and replicate within host cells, with the bacterial cell outer membrane (OM) playing a key role in this process. The Brucella OM contains three major proteins, Omp25, Omp31, and Omp2b,2 with molecular masses of 25–27 kDa, 31–34 kDa, and 36–38 kDa, respectively. Omp25 stimulates both cell-mediated and humoral immunity in humans and plays an important role in the secretion of TNF-α and IFN-α in macrophage and cytokine cells.3–5
To develop efficient vaccines against brucellosis, the relationship between Omp25 and virulence has been the focus of much research. However, the results have been contradictory. Edmonds et al. reported that the virulence of B. melitensis and B. ovis omp25-deficient mutants is attenuated,6 while Manterola et al. showed that the absence of omp25 does not result in attenuation of virulence of B. abortus.7 These findings suggest that the interaction between Omp25 and host cells is very complex.
MicroRNAs (miRNAs) are noncoding RNAs approximately 22 bp in length that function as a sequence-specific, post-transcriptional regulatory mechanism.8 Multiple reports indicate that miRNAs play important roles in the Brucella infection process. For example, miR-125b-5p is down-regulated during B. abortus infection, which enhances the expression of its target gene, A20. In addition, B. abortus-induced down-regulation of miR-125b-5p is linked to enhanced intracellular growth of the pathogen via regulation of macrophage activation.9 In another study, miRNA array and quantitative real-time PCR (qRT-PCR) analyses were performed using peripheral blood mononuclear cell samples from patients with acute or chronic brucellosis to identify differentially expressed miRNAs. The assays identified four miRNAs expressed in the chronic group that were not expressed in the acute and control groups. Among these, the expression of miR-1238-3p was increased while that of miR-494, miR-6069, and miR-139-3p was decreased.10 Further, an array-based miRNA screen using CD8+ T-cells from patients with acute or chronic brucellosis indicated that the differentially expressed miRNAs miR-126-5p and miR-4753-3p, along with their predicted target genes, are involved in the MAPK signaling pathway, cytokine–cytokine receptor interactions, endocytosis, regulation of the actin cytoskeleton, and focal adhesion.11
In the current study, we constructed an Omp25-deficient B. melitensis strain (M5-90-Δomp25) to identify functional molecules in macrophages, including miRNAs and their target genes, which are affected by Brucella Omp25. miRNA and mRNA profiling conducted in RAW264.7 cells infected with B. melitensis M5-90-Δomp25 identified four differentially expressed miRNAs along with 10 potential target genes that are differentially expressed resulting of their interaction with Omp25.
Materials and methods
Cells and bacterial infection
RAW264.7 macrophages were grown at 37℃ and 5% CO2 in DMEM (Invitrogen, Carlsbad, CA). B. melitensis strain M5-90 was grown in Rubella broth medium (BD Biosciences, Franklin Lakes, NJ) for 3 d at 37℃ prior to use. For infection studies, RAW264.7 cells at a density of 1 × 106 cells/well (in six-well plates) were infected with the bacterial inoculums at a multiplicity of infection of 10 and incubated for 4 h. Following infection, cells were washed three times with phosphate-buffered saline (PBS) to remove extracellular bacteria.12
Preparation of the omp25 B. melitensis deletion mutant and its use in infection assays
Three pairs of primers, Omp25-C-F/Omp25-C-R and Omp25-N-F/Omp25-N-R and Kanar-F/Kanar-R (Table 1), were designed to amplify the flanking regions of the omp25 gene from the genome of B. melitensis M5-90 as well as a kanamycin-resistance gene fragment from pEGFP-N1, respectively. The three fragments were separately ligated into vector pMD20-T, generating recombinant plasmids pMD20-T-Omp25-C, pMD20-T-Omp25-N, and pMD20-T-Kanar. pMD20-T-Omp25-C was then digested with SacI and BamHI, pMD20-T-Kanar with BamHI and KpnI, and pMD20-T-Omp25-N with KpnI and ApaI. The pGEM-7ZF suicide plasmid was also digested with the same enzymes to produce corresponding sticky ends. The digested suicide plasmid was then connected with the omp25 upstream region, the Kanar gene fragment, and the omp25 downstream region in a stepwise manner and then confirmed by sequencing. The correct recombinant plasmid was then transformed into electrocompetent B. melitensis M5-90 cells and recombinants were selected on medium containing kanamycin and ampicillin. PCR amplification using the Kanar-F and Kanar-R primers was performed to confirm the recombination events. The obtained strain was named B. melitensis M5-90-Δomp25. For infection assays, RAW264.7 cells were infected with either B. melitensis M5-90 or B. melitensis M5-90-Δomp25 cells and incubated for 4 h, as described previously.12
Primers for constructing B. melitensis M5-90-ΔOMP25.
Primers
Sequences (5′-3′)
Inserted enzyme
OMP25-C-F
GCGCGAGCTCTTATAGCATAATTGGACACG
Sac I
OMP25-C-R
CGCGGGATCCAAAAGTGAAT ATTCCGCC
BamH I
Kanr-F
GCGCCCCGGGATGATTGAACAAGATGGATT
BamH I
Kanr-R
CGCGCTCGAGTCAGAAGAACTCGTCAAG
Kpn I
OMP25-N-F
GCGCGGTACCCTAACCTGCCGCTCGAAA
Kpn I
OMP25-N-R
CGCGGGGCCCGGCATTCTCCTTACACAAAT
Apa I
OMP25-F
CGCGGAATTCATGGCGCACTCTTAAGTCTC
EcoR I
OMP25-R
GCGCCTCGAGGAACTTGTAGCCGATGCC
Xho I
OMP25-C-J-F
CGTGATATTGCTGAAGAGC
OMP25-C-J–R
ATTACCTTCAGGAAGTCCG
OMP25-N-J-F
TCGCGTTCCACAAAGCCG
OMP25-N-J-R
GGACAGGTCGGTCTTGACAA
Ampr-F
ATGAGTATTCAACATTTCCG
Ampr-R
TTACCAATGCTTAATCAGTG
miRNA array screening and qRT-PCR validation
miRNA was extracted from uninfected RAW264.7 cells and from RAW264.7 cells infected with either B. melitensis M5-90 or B. melitensis M5-90-Δomp25 as described previously.13 miRNA microarray analyses were carried out by LC Sciences (Houston, TX). Using miRNA-specific primers obtained from Invitrogen, qRT-PCR analysis was performed to validate the differentially expressed miRNAs identified in the array-based screen, as described previously.12,13 U6 snRNA was used as an internal control. Each assay from different RNA sample was repeated three times.
mRNA array screening
High-quality total RNA was extracted from RAW264.7 cells infected with B. melitensis M5-90 or B. melitensis M5-90-Δomp25 as described previously.12 Gene expression profiling and analysis was performed using the Agilent platform (V2, G4846A/26655) by LC-BIO Co. (China). Significantly differentially expressed genes (DEGs) (fold change ≥2) were selected for further miRNA–mRNA conjoint analysis.
miRNA–mRNA conjoint analysis
TargetScan, miRanda, and PicTar were used to predict the target genes of the validated differentially expressed miRNAs as previously described.13 Of the potential target genes predicted by TargetScan, those with a context, score, and percentile lower than 50 were removed. Using miRanda, potential target genes with a Max Energy higher than −10 were removed, while potential target genes predicted using PicTar with a dideoxyguanosine score higher than −5 were also removed. The three lists were then collated, and genes appearing on all three lists were selected as the predicted target genes of the validated differentially expressed miRNAs.
Further comparison of the prediction-based gene list and the previous array-based gene list was performed, and the overlapping target genes were further validated by qRT-PCR analysis.
qRT-PCR validation of miRNA–mRNA conjoint analysis results
qRT-PCR was performed to confirm the DEGs obtained from the miRNA–mRNA conjoint analysis as previously described,12,13 using the specific primers described in Table 2. GAPDH was used as an internal control. Each RNA sample was examined in triplicate.
Primers of qRT-PCR identification for the DEGs (fold change>2) obtained from miRNAs–mRNA conjoint analysis.
Gene symbol
GenBank ID
Primer sequence (5′-3′)
Bcl6b
NM_007528
F: AAGAGCGCACAAGGCAGTTC
R: CCCCGGAAAATTGAATAGAAGA
Prss46
NM_183103
F: CCCCTCGTCCAGCCAATC
R: TGGTTCCAATGCTTGGCTTT
Olr1
NM_138648
F: CTTCCATGGGCCCTTTAGC
R: TGGCCACCCAAAGATTGG
Ikbke
NM_019777
F: CGGTCATCCACGTCTTTTCC
R: GGGCGTGGATGTAGACATGAT
Ceacam14
NM_025957
F: TGCTCACAGTCTCCCTTTTAACC
R: TTTGATGGTGAGCTGGGAAGT
Slc7a11
NM_011990
F: GCACCTAAGCTCCTTCCAAGAC
R: GATAGCTGCCATATGTATTCTCTGTAAAG
Asxl3
NM_001167777
F: TCTGAATGCCATGCTTCACACT
R: CAACGACTAGATCCAATGTTCCAT
Mras
NM_008624
F: CAATGCCAGATACTTGAAACAATGA
R: CGCTCCTCACACGACAACTG
Slc31a2
NM_025286
F: TTGAGATGAAACCAGCAAATGG
R: GGGAGGGAGGTAGCTTCAATG
Cdk1
NM_007659
F: GGACGAGAACGGCTTGGAT
R: GAGATTCGTTTGGCAGGATCA
Il1rl1
NM_001025602
F: ACAGACAGAGAATGGGACTTTGG
R: ACTGCCCTCCGTAACTGTCAA
Nos2
NM_010927
F: GCCCCACGGGACACAGT
R: AACAGCTCAGTCCCTTCACCAA
Ifit1
NM_008331
F: GGCAGGTTTCTGAGGAGTTCTG
R: TCCCATGGTTGCTGTAAAGGT
Dusp16
NM_130447
F: TCCTGGCTTGAGTTGTATCCTCTA
R: CCCTGCTGACGTGAGTGAATT
Obox6
NM_145710
F: GCAACCCTCTGATTTCAGGTTCT
R: GGCATATGTGGGCTCTGATTG
Tbr1
NM_009322
F: GATCCCAATCACTGGAGGTTTC
R: GTGTCCGCTTTGCCACAAG
Igfbp2
NM_008342
F: GCCCCCTGGAACATCTCTACT
R: TACCGGCCATGCTTGTCA
GAPDH
NM_008084.3
F:TGTGTCCGTCGTGGATCTGA R:CCTGCTTCACCACCCTTCTTGA
Statistical analysis
A Student’s t-test or one-way ANOVA was used to compare the data from the different treatment groups. P < 0.05 was considered statistically significant. P < 0.01 was considered very statistically significant.
Results
Differentially expressed miRNAs in B. melitensis M5-90-Δomp25- infected RAW264.7 cells
RAW264.7 cells were infected with B. melitensis M5-90 or B. melitensis M5-90-Δomp25 and then examined after incubation for 4 h. Initial microarray-based screening identified eight differentially expressed miRNAs (Figure 1 and Table 3). Of these, the expression of mmu-miR-146a-5p, mmu-miR-155-5p, and mmu-miR-3473a was up-regulated, while expression of mmu-miR-149-3p, mmu-miR-671-5p, mmu-miR-1224-5p, mmu-miR-1895, and mmu-miR-5126 was down-regulated. The results of further qRT-PCR validation experiments confirmed that mmu-miR-146-a-5p and mmu-miR-155-5p were up-regulated and that mmu-miR-149-3p and mmu-miR-5126 were down-regulated (Figure 2), and that the strategy of initial array-based screening and further qRT-PCR validation was feasible. However, four of the initial differentially expressed miRNAs were not confirmed by qRT-PCR, suggesting the presence of false-positive signals in the array-based screen.
Heat map of differentially expressed miRNAs in RAW264.7 infected with Omp25 mutant B. melitensis (fold change >2) (P < 0.01). The experiments in which RAW264.7 were infected with B. melitensis M5-90 was repeated three times. M5-90-1, M5-90-2, and M5-90-3 deciphers the sample of each infection assay, respectively. The experiments in which RAW264.7 were infected with B. melitensis M5-90 ΔOmp25 was repeated three times. ΔOmp25-1, ΔOmp25-2, and ΔOmp25-3 deciphers the sample of each infection assay, respectively. Red denotes induction and green denotes suppression.
qRT-PCR validation of the differentially expressed miRNAs obtained from the array-based intial screening.
Significantly differentially expressed miRNAs.
Group1 M5-90
Group2 ΔOMP25
Log2
Reporter name
P-value
Mean
Mean
(G2/G1)
mmu-miR-146a-5p
3.10E-03
877
2,056
1.23
mmu-miR-155-5p
1.69E-03
388
1,677
2.11
mmu-miR-3473a
9.06E-03
77
119
0.63
mmu-miR-149-3p
6.90E-03
540
215
−1.33
mmu-miR-671-5p
7.19E-03
39
22
−0.87
mmu-miR-1224-5p
5.49E-03
155
46
−1.76
mmu-miR-1895
5.36E-03
207
110
−0.91
mmu-miR-5126
2.41E-03
1,214
296
−2.04
Differentially expressed mRNAs in B. melitensis M5-90-Δomp25-infected RAW264.7 cells
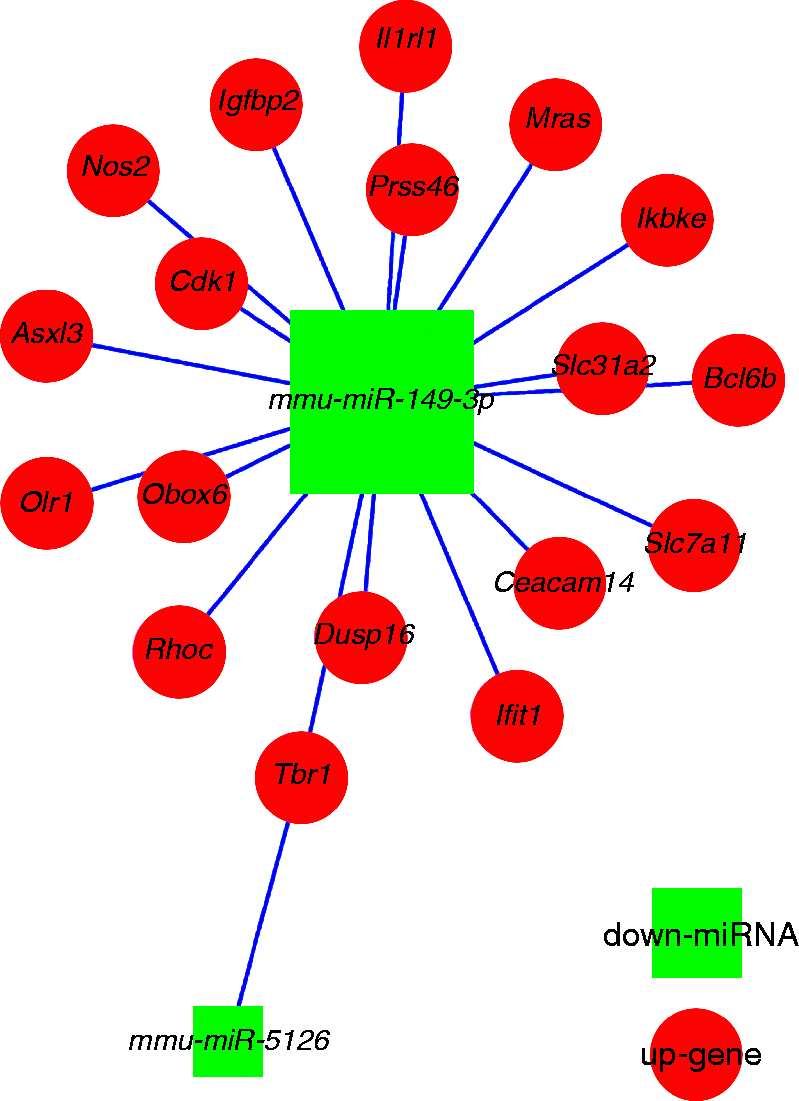
As shown in Figure 3 and Supplementary Figure 1, the mRNA microarray identified 967 DEGs in RAW264.7 cells infected with B. melitensis M5-90-Δomp25. Further miRNA-mRNA conjoint analysis indicated that among the 967 DEGs, 17 genes, namely Slc7a119 (also known as xCT), Mras (muscle RAS oncogene homolog), Slc31a2 (solute carrier family 31 member 2), Asxl3 (additional sex combs-like 3), Ifit1 (interferon-induced with tetratricopeptide repeats), Obox6 (oocyte-specific homeobox 6), Cdk1 (cyclin-dependent kinase 1), Ceacam14 (CEA-related cell adhesion molecule14), Dusp16 (dual-specificity phosphatase 16), Igfbp2 (insulin-like growth factor-binding protein 2), Ikbke (inhibitor of kappaB kinase epsilon), Olr1 (oxidized low density lipoprotein (lectin-like) receptor 1), Prss46 (proteases, serine), Tbr1 (T-box brain 1), Bcl6b (B cell CLL/lymphoma 6, member B), Il1rl1 (IL-1 receptor-like 1), and Nos2 (NO synthase 2, inducible), were predicted to be target genes of mmu-miR-149-3p by the bioinformatics analyses, and were up-regulated. One of these 17 genes, Tbr1, was also predicted to be a target of two further miRNAs, mmu-miR-149-3p and mmu-miR-5126, and to be up-regulated in both cases (Table 4, Figures 4 and 5).
Statistical graph of the numbers of DEGs obtained from the array-based intial screening in RAW264.7 infected with Omp25 mutant B. melitensis.
The validated differentially expressed miRNAs and their predicted targeting genes in RAW264.7 infected with Omp25 mutant B. melitensis.
Heat map of the DEGs selected from mRNA array screening in RAW264.7 infected with Omp25 mutant B. melitensis (fold change ≥ 2) (P < 0.05). These DEGs are also the predicted targeting genes of the validated differentially expressed miRNAs. The experiments in which RAW264.7 were infected with B. melitensis M5-90 was repeated three times. M5-90-1, M5-90-2, and M5-90-3 was the sample of each infection assay, respectively. The experiments in which RAW264.7 were infected with B. melitensis M5-90 ΔOmp25 was repeated three times. ΔOmp25-1, ΔOmp25-2, and ΔOmp25-3 was the sample of each infection assay, respectively. Red denotes induction and green denotes suppression.
mRNA array-based expression levels of DEGs (fold change>2) obtained from miRNAs-mRNA conjoint analysis.
Group1 M5-90
Group2 ΔOmp25
Log2
Gene symbol
P-value
Mean
Mean
(G2/G1)
Bcl6b
1.85E-02
27.61
74.78
1.44
Prss46
3.61E-02
84.56
185.70
1.13
Olr1
3.83E-02
61.37
137.79
1.17
Ikbke
3.69E-03
54.41
135.77
1.32
Ceacam14
1.49E-03
20.08
50.91
1.34
Slc7a11
8.17E-03
35.95
161.74
2.17
Asxl3
8.10E-03
6.39
20.20
1.66
Mras
1.49E-02
25.39
75.84
1.58
Slc31a2
9.61E-03
271.65
991.51
1.87
Cdk1
3.13E-03
47.74
115.75
1.28
Il1rl1
1.75E-04
642.66
1461.13
1.18
Nos2
1.31E-03
114.00
869.50
2.93
Ifit1
2.94E-03
545.90
1548.83
1.50
Dusp16
2.82E-03
253.67
627.60
1.31
Obox6
2.87E-03
6.39
20.75
1.70
Tbr1
3.78E-02
26.10
71.27
1.45
Igfbp2
1.58E-02
19.69
39.67
1.01
After specific primers were designed (Table 2), qRT-PCR validation of the predicted target genes was performed. The results confirmed the up-regulation of Bcl6b, Nos2, Ikbke, Slc31a2, Dusp16, Ifit1, Slc7a11, Il1rl1, and Olr1 (Figure 6).
qRT-PCR validation for the identified up-regulated DEGs in RAW264.7-M-Δ Omp25.
Discussion
Brucella species are Gram-negative, intracellular bacteria responsible for the zoonosis brucellosis. To survive within host cells, Brucella species affect the secretion of cytokines, including TNF-α, and induce dysregulation of the normal protective function of the immune response.3,14 During the infection process, Omp25 on the surface of Brucella cells controls TNF-α production in human macrophages.15 Functional insight into Omp25 would aid in elucidating the virulence mechanisms of Brucella. B. melitensis vaccine strain M5-90 has some residual virulence and may result in abortion in pregnant sheep. In the current study, we constructed Omp25-deficient strain B. melitensis M5-90-Δomp25 to examine the function of Omp25.
To improve the signal/noise ratio when identifying DEGs in RAW264.7 cells infected with B. melitensis M5-90-Δomp25, we used a research strategy that generated two gene lists from different analysis methods. After differentially expressed miRNAs were identified, their target genes were predicted using different bioinformatics software tools, and these target genes formed the first gene list. The second gene list was obtained from mRNA array-based screening. The overlapping genes from the two gene lists formed the final list of target genes. qRT-PCR analysis was performed to validate the DEGs on the final list. Among these overlapping genes, nine genes, all of which are targets of mmu-miR-149-3p, were identified as the DEGs of RAW264.7 cells infected with B. melitensis M5-90-Δomp25. The results of qRT-PCR validation demonstrated that the strategy is feasible and effective.
Bcl6b, Nos2, Ikbke, Slc31a2, Dusp16, Ifit1, Slc7a11, Il1rl1, and Olr1 were confirmed as the target genes of mmu-miR-149-3p. In humans, BCL6b is expressed in a small subset of Ag-experienced CD8+ T-cells and is required for the enhanced secondary response of memory CD8+ T-cells,16 while NOS2 is involved in the production of NO, a cellular defense mechanism against Brucella infection.17 DUSPs are a family of protein phosphatases.18 In macrophages, Dusp16 RNA expression is inducible by TLR stimulation.19,20 Dusp16 preferentially dephosphorylates c-Jun N-terminal kinase- and p38-family MAPKs.21 In infected macrophages, to suppress host immune responses for intracellular survival, Mycobacterium tuberculosis Eis (enhanced intracellular survival protein) initiates the inhibition of autophagy and phagosome maturation by Nɛ-acetylating Lys55 of DUSP16.22 IFIT proteins are critical mediators of mammalian innate antiviral immunity, with IFIT1 proving to be a potent inhibitor of alphavirus replication.23 In the mouse lung, intraperitoneal challenge with LPS induces the up-regulation of Olr1 expression, suggesting that Olr1 is involved in LPS-induced lung inflammation.24 The up-regulation of all of these genes in our infection model indicates that the functional mechanism of Omp25 might involve multiple signaling pathways and a large number of signaling molecules.
This is the first report describing miRNA expression analysis of macrophage cells infected with an Omp25-deficient Brucella strain. Although further efforts are needed to fully elucidate the interaction between Omp25 and these molecules, our findings provide some important information that will aid in the prophylaxis and treatment of brucellosis.
Supplemental Material
Supplemental material for Alterations of microRNAs and their predicted targeting mRNAs expression in RAW264.7 macrophages infected with Omp25 mutant Brucella melitensis
Supplemental material for Alterations of microRNAs and their predicted targeting mRNAs expression in RAW264.7 macrophages infected with Omp25 mutant Brucella melitensis by Huapei Zhu, Hanwei Jiao, Xin Nie, Baobao Li, Kailian Xu, Feng Pang, Ruiyong Cao, Shu Zhu, Xiaojian Yang, Zhenxing Zhang, Dongmei Peng, Yaying Li, Guohua Li, Haifeng Huang, Chuangfu Chen, Li Du and Fengyang Wang in Innate Immunity
Footnotes
Declaration of conflicting interests
The author(s) declare no potential conflicts of interest with respect to the research,authorship,and/or publication of this article.
Funding
The author(s) disclose receipt of the following financial support for the research,authorship,and/or publication of this article: This work was financially supported by the Key Science and Technology Project of Hainan (ZDKJ2016017-01),the Key Research and Development Program of China (No. 2017YFD0500302),the China Agriculture Research System (CARS-38),and the Natural Science Foundation of China (31460670).
CloeckaertAVizcainoNPaquetJYet al.Major outer membrane proteins of Brucella spp.: past, present and future. Vet Microbiol2002; 90: 229–247.
3.
DornandJGrossALafontVet al.The innate immune response against Brucella in humans. Vet Microbiol2002; 90: 383–394.
4.
MaQLLiuACMaXJet al.Brucella outer membrane protein Omp25 induces microglial cells in vitro to secrete inflammatory cytokines and inhibit apoptosis. Int J Clin Exp Med2015; 8: 17530–17535.
5.
SalhiIBoigegrainRAMacholdJet al.Characterization of new members of the group 3 outer membrane protein family of Brucella spp. Infection and Immunity2003; 71: 4326–4332.
6.
EdmondsMDCloeckaertAElzerPH. Brucella species lacking the major outer membrane protein Omp25 are attenuated in mice and protect against Brucella melitensis and Brucella ovis. Vet Microbiol2002; 88: 205–221.
7.
ManterolaLGuzman-VerriCChaves-OlarteEet al.BvrR/BvrS-controlled outer membrane proteins Omp3a and Omp3b are not essential for Brucella abortus virulence. Infection and Immunity2007; 75: 4867–4874.
8.
Lagos-QuintanaMRauhutRLendeckelWet al.Identification of novel genes coding for small expressed RNAs. Science2001; 294: 853–858.
9.
LiuNWangLSunCet al.MicroRNA-125b-5p suppresses Brucella abortus intracellular survival via control of A20 expression. BMC Microbiol2016; 16: 171–171.
10.
BudakFBalSHTezcanGet al.Altered expressions of miR-1238-3p, miR-494, miR-6069, and miR-139-3p in the formation of chronic brucellosis. J Immunol Res2016; 2016: 4591468–4591468.
11.
BudakFBalSHTezcanGet al.MicroRNA expression patterns of CD8+ T cells in acute and chronic brucellosis. PLoS One2016; 11: e0165138–e0165138.
12.
LeiMDuLJiaoHWet al.Inhibition of mCD14 inhibits TNF-α secretion and al. NO production in RAW264.7 cells stimulated by Brucella melitensis infection. Vet Microbiol2012; 160: 362–368.
13.
ChengYKuangWHHaoYCet al.Downregulation of miR-27a* and miR-532-5p and upregulation of miR-146a and miR-155 in LPS-induced RAW264.7 macrophage cells. Inflammation2012; 35: 1308–1313.
14.
ChaSBRayamajhiNLeeWJet al.Generation and envelope protein analysis of internalization defective Brucella abortus mutants in professional phagocytes, RAW 264.7. FEMS Immunol Med Microbiol2012; 64: 244–254.
15.
Jubier-MaurinVBoigegrainRACloeckaertAet al.Major outer membrane protein Omp25 of Brucella suis is involved in inhibition of tumor necrosis factor alpha production during infection of human macrophages. Infect Immun2001; 69: 4823–4830.
16.
MandersPMHunterPJTelarantaAIet al.BCL6b mediates the enhanced magnitude of the secondary response of memory CD8+ T lymphocytes. Proc Natl Acad Sci USA2005; 102: 7418–7425.
17.
WangMQureshiNSoeurtNet al.High levels of nitric oxide production decrease early but increase late survival of Brucella abortus in macrophages. Microb Pathog2001; 31: 221–230.
18.
ZhangHZhengHMuWet al.DUSP16 ablation arrests the cell cycle and induces cellular senescence. FEBS J2015; 282: 4580–4594.
19.
MatsuguchiTMusikacharoenTJohnsonTRet al.A novel mitogen-activated protein kinase phosphatase is an important negative regulator of lipopolysaccharide-mediated c-Jun N-terminal kinase activation in mouse macrophage cell lines. Mol Cell Biol2001; 21: 6999–7009.
20.
NiedzielskaMBodendorferBMünchSet al.Gene trap mice reveal an essential function of dual specificity phosphatase Dusp16/MKP-7 in perinatal survival and regulation of Toll-like receptor (TLR)-induced cytokine production. J Biol Chem2014; 289: 2112–2126.
21.
TanoueTYamamotoTMaedaRet al.A novel MAPK phosphatase MKP-7 acts preferentially on JNK/SAPK and p38α and β MAPKs. J Biol Chem2001; 276: 26629–26639.
22.
KimKHAnDRSongJet al.Mycobacterium tuberculosis Eis protein initiates suppression of host immune responses by acetylation of DUSP16/MKP-7. Proc Natl Acad Sci USA2012; 109: 7729–7734.
23.
ReynaudJMKimDYAtashevaSet al.IFIT1 differentially interferes with translation and replication of alphavirus genomes and promotes induction of type I interferon. PLoS Pathog2015; 11: e1004863–e1004863.
24.
ZhangPLiuMCChengLet al.Blockade of LOX-1 prevents endotoxin-induced acute lung inflammation and injury in mice. J Innate Immun2009; 1: 358–365.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.