
Abstract
1. Introduction
Fish have a number of swimming modes that are worth considering for emulation, but the locomotion that has long attracted the attentions of both biologists [1] and engineers [2]–[5] is the Body/Caudal fin undulatory kinematics. Many papers have addressed the live undulatory locomotion of fish over the past few years. But studying how fish generate the external fluid force to swim has proven to be difficult and is limited in the ability of controlling for parameters in a precise and repeatable manner. There still exists several important questions about kinematic effects on the hydrodynamics [6]. In many previous studies of fish swimming, the fish undulatory body and flapping caudal fin kinematics are treated together as a derivable mathematical wave [7][8]. Nevertheless, recent biological findings indicate that the caudal fin undergoes complex kinematics independent of the body in lots of bony fishes [6]. Both body undulation and caudal fin flapping play essential roles while a fish is swimming. How do these two parts inference the hydrodynamics of the fish individually? To what extent does each part of these two contribute to the over-all swimming performance? In addition, how is the wake flow generated as a function of the precisely controlled kinematics of the undulatory body and the flapping caudal fin? The questions presented above are certainly not the only possible ones worth discussion, but can serve as a starting point for thinking about.
Understanding how different locomotion of swimmers affects their thrust performance requires qualitative hydrodynamic experiments [9]. Simultaneous measurements of the hydrodynamic quantities (the fluid power, thrust force and self-propulsive speed) are strongly recommended [10]. The promise of robotic models for studying the biomechanics of locomotion in fishes has just begun to be realized. The robotic model offers a complementary approach to studies of living organisms by allowing precise control of the motion, accurate measurements of the forces and the exploration of a range of kinematic parameter spaces broader than that which exists in nature, also including kinematics that are not commonly used by live swimmers [10]. Both biologists and Engineers are particularly interested in how robotic swimmers quantitatively interact with their surrounding fluid environment. Through many studies on fish biomechanics [9], one significant technological development stands out. By seeding particles in water and shooting a laser sheet behind the swimming fish or robotic model, we were able to quantify hydrodynamic variables and flow patterns directly by using Digital Particle Image Velocimetry (DPIV) [11]–[15].
The quantitative thrust efficiency (t) and self-propulsive swimming speeds (
2. Materials and methods
A. Experimental apparatus and self-propulsion method
Fig.1 shows the schematic overview of the present experimental apparatus for the hydrodynamic experiment. The servo towing system is driven by a 4,000 watt AC motor and has a travel distance of 7.5m. Underneath the towing system is a water tank measuring 7.8mx1.2mx1.1m. The robotic fish moves at mid-depth in the tank to avoid the interference effect of the free surface and the bottom of the tank. As Fig.1a shows, the robotic fish and its mechanical transmission part are fixed under the force transducer. A low-drag streamlined strut penetrates the water and connects with the head of the fish. The external force of a robotic fish can be measured using a multi-component piezoelectric force transducer (Kislter 9254C). The output of the external Kislter transducer is recorded by Computer I through a data cable using a Controller Area Network (CAN) bus.
In the present experimental method, the self-propulsive speed is applied under a constant towing speed
The DPIV system, which was fixed on the carriage (as can be seen in the schematic view), was used to measure the flow field generated by the robotic fish. A high speed CCD camera (100 fps, 1024 pixels × 1024 pixels, Mintron Inc.) with an optical band pass filter is used to capture the particle image of the flow field. The laser beam (4W) with a wavelength of 0.532 μm is expanded by two cylindrical lenses to generate a light sheet that is reflected successively by three mirrors into the water tank and passes through the midline of the caudal fin of the fish. The wake is visualized by seeding the water with nylon particles (40–70 μm, 1.05 g cm3) to reflect the laser. This resulted in the successful illumination of the captured area by the bright laser light sheet. We then used the ‘mpiv’ Matlab toolbox, which is open-source software and was successfully tested in our previous biological studies [15]. As shown in the dashed blue box in Fig. 1a, when the experimental apparatus was in operation we were able to simultaneously record the power measurement system, DPIV system, force transducers that were mounted on the carriage running in the axial direction with speed (denoted by

(a) Schematic overview of the experimental apparatus. Under condition of self-propulsion, the time-average thrust force and drag force are balanced. (b) Snapshot of the experimental robotic model with laser system working. (c) High-speed camera recorded image. The white dashed box indicates the captured area of the particle images and the upward and downward flick indicates the side-to-side movement in reality.
B. The Biomimetic robotic fish and kinematics
The mackerel (

(a). Simulation of the robotic links in ADAMS. (b) Angles for different discrete segments.

(a) (b) Robotic fish body undulation at instants of
To study the hydrodynamic effect of the robotic swimmer as a function of the Stouhal number (


Where
Fig. 3a and 3b show snapshots of the undulating robotic fish in two instances. We digitalized the midline of the robotic fish body from a bottom view in Matlab (the robotic fish is upside-down). Fig.3c and 3d show the midline curves of imposing programmed kinematics on the robotic mackerel's body. Note that only the posterior 2/3 of body moves. Midlines at equally spaced time intervals throughout a tail beat can be observed. Each time this is shown in a distinct colour. The movement of the robotic fish is quite relevant to the kinematics of the live swimmer (more live fish body midline images can be referred to Fig. 11.1 on page 427 of [1]).

(a) Schematic views of single-row and double-row vortices. (b) Three dimensional vortex model and laser sheet. Different colours indicate the distinct directions of vortex rotation, and the arrow indicates the direction of the jet flow.
According to the study of Tytell [18], the wavelength of typical anguilliform kinematics is about 0.65
Parameters of undulation kinematics

From Table 1, systematic tests were also performed for the Strouhal number, which were varied in small increments from 0.15 to 1.05. This
Previous experimental studies of the flapping foil [19] and the swimming live fish [9] show that the hydrodynamic performance of fish locomotion is dominated by the fundamental dimensionless parameter (the Strouhal number), which is defined as:
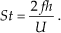
where

where |·|max denotes the maximum absolute value. The Reynolds number (

where

where ρ is the density of the fluid and


where σ is the sample standard deviation for each data point consisting of
C. DPIV and thrust force estimation
To measure the flow field and then calculate the thrust force of the swimming robotic fish using a two-dimensional approach, the laser sheet plane of the DPIV system should be set to pass through the midline of the Homocercal caudal fin of the robotic fish. The midline was recommended as the best horizontal position for conducting two-dimensional wake measurements [9]. The depth of the three-dimensional vortex rings (see Fig. 3a for notation) effectively equals the caudal fin's height, while the measurement planes at other horizontal levels underestimate the thrust force.
The vortex ring model, which assumes that all the impulse shed by the robotic fish is contained in elliptical vortex rings, is used for the analysis of the wake [24]. The measured wake then allows us to use 2D vortex dynamics theory to compute the impulse of the fish. We form the flow field from several tail flap cycles by freezing each vortex in its shed position and then take measurements of the wake. There should in fact be two different types of wake structures generated by the robotic swimmer. This point was confirmed in the current experiment. In Fig. 3b, the distribution and morphology of the vortices are provided in a three-dimensional way.
Illuminating a cross-section through the three-dimensional vortex ring yields a flow pattern consisting of two vortices. The location of the vortices in the velocity fields was determined by plotting the contours of vorticity. The morphology of a vortex is described by the vortex centre, the core radius

where

where

where α1 and α2 denote the jet angle for two vortex rings. Details of the calculation of thrust force using the vortex dynamic model can be referred to in our previous study on live fish. The thrust efficiency is defined following Lighthill [23]:

3. Experimental Results
A. Self-propulsive speed result
From Fig. 4a it can be seen that as the

(a) Strouhal number (
The point at which the
Recalling previous biological studies (Hess and Videler, 1984), the live mackerel vary their flapping amplitude
B. DPIV Flow field and wake structure
Fig. 5 shows the DPIV time-series of the flow field that is generated by the robotic swimmer at

Flow field of the robotic swimmer with kinematics parameters of
Figure 6(a) is a main schematic depiction of the wake generated by the current robotic model and summarizes our experimental findings of wake geometry. By varying the kinematic parameters we visualize a variety of wake structures, including the Von Karman structure, Single-row Reverse Karman wake (2S), Two-Paired vortices (2P) wake and Two-Paired and Two-Single vortices (2P+2S) wakes from the characteristic of the near wake flow. Present results showed that the near flow wake structure is less sensitive to the wavelength

(a) The DPIV results of different wake structures generated by the robotic model. Panel A∼D show the vortices and flow vectors at the same instance of one tail beat, where
To explain the general trend in wake transition observed in this study, the lowest
A very interesting result can be observed. Within the range of 0.325≤
Nevertheless, present DPIV results showed that both the caudal fin pitch angle and the Strouhal number had a significant impact on the wake structure. Additionally, we found a Two-Paired and Two-Single vortices (2P+2S) wake structure in region D, also from the characteristic of the near wake flow. Despite the fact that some previous studies on flapping foils showed the wake transition [25], those experiments are not conducted under the condition of self-propulsion. Also, the isolated performance of a flapping foil cannot fully represent a swimming fish's, especially the body/caudal fin kinetic pattern that characterizes approximately 88% of extant fish families [8].
C. Thrust efficiency
Fig. 7 shows the estimated thrust efficiency η as a function of the

Thrust efficiency as function of Strouhal number
Fig 8a demonstrates that the peak thrust efficiency was 38.1%, recorded at 0=10°. The thrust efficiency decreased as 0 increased within the range of 25°<θ<40°. At 0=40° the thrust efficiency reduced to a minimal value of 13.6%. It is quite interesting that the peak self-propulsion Strouhal number (

(a) Thrust efficiency and Self-propulsion
4. Discussion
To our knowledge, there exist very limited publications that report the quantitative hydrodynamic performance of the freely swimming robotic fish. The only relevant study is conducted by the Techet group [26], who applied DPIV to characterize the swimming performance of a complaint fish-like robot. The midline kinematics result of the robot showed that the undulatory body and the caudal fin formed a derivable mathematical curve. The drawback of this work is that the robot is made of a continuous flexible body so the body and caudal fin cannot be individually controlled. The minimal
We also showed that the wake structure of the robotic model depend strongly on the Strouhal number and caudal fin pitch angle. For robotic fish swimming in a self-propelled condition, the caudal fin pitch angle proved to be an essential factor governing the transition of the wake structure and the thrust efficiency. The body wavelength, which also affects the thrust efficiency, however, does not have a significant effect on the wake structure. The reverse Karman wake is produced by many fish species and physical systems, such as the oscillating thrust-producing foil moving steadily forward [19] and during the caudal fin locomotion by bluegill sunfish (
Note that, in the current study, we use a two-dimensional planar technique at the mid-span of caudal fin for investigating a variety of principal parameters relating to undulatory propulsion in fishes. Present data would be difficult to obtain directly from studying live fishes. The wake flow in aquatic locomotion appears to be affected by a three-dimensional fluid effect. Three-dimensional PIV techniques, such as volumetric imaging, should be applied to understand the fluid physics of fish locomotion, which would possibly reveal new features of fish locomotor dynamics.
5. Conclusion
Robotic techniques are becoming increasingly important in the field of biomechanics [27]. They offer the opportunity to focus research by creating robotic models that cannot be easily controlled with the desired kinetic mechanisms. In this paper, we simultaneously measured the power consumption, self-propulsion speed and wake structure of a robotic model. Results show that the wake structure is more sensitive to the caudal fin pitch angle (